私が共著者に加わっている論文がプレプリントサーバーのBioRvixで公開されました。プレプリントサーバーは受理されていない段階の論文を公開する仕組みで、生態学ではまだ少ないようですが、科学界全体では多くの論文がプレプリントサーバーで公開されるようになってきています。
論文査読の仕組みとプレプリントサーバーの登場
論文が学術誌に掲載されるまでには「査読」というプロセスがあります。研究者の間では査読がされないレポートなどと区別するために「査読論文」という呼び方が普通に用いられているほどで、査読を受けているか否かは研究の信憑性を判断するための重要な基準になっています。査読ではその分野の専門家(一般的には2名)が論文の草稿を読んで問題点を指摘します。査読を通過することを「受理」と言い、受理されると論文は研究業績として認められたことになります。そして受理された論文が雑誌やWebサイトに公開されることを「出版」と言います。
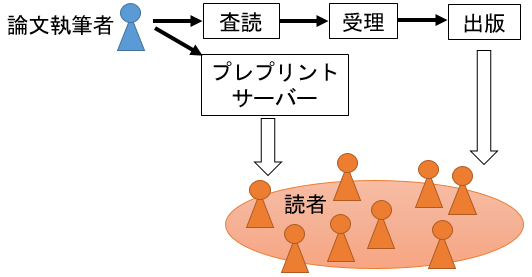
図1. 論文が公開される流れ。従来は査読を経て出版されるまで内容を読むことができなかったが、プレプリントサーバーで公開までの時間が短縮された。ただし査読で審査されていないため、誤りがあったり、出版までに内容が変わることもあり得る。
研究者が論文を書いて学術誌に送ると、まず編集者が内容をチェックし、その学術誌に掲載する価値がある内容だと認められなければこの段階で却下され(学術誌にはそのレベルにふさわしい内容が求められます)、価値ありと判断されると査読が始まります。執筆者は査読者の指摘に応じて何度も論文を修正しますが、査読者に納得してもらえなければ却下になります。そうなると、もう少し難易度の低い学術誌に再投稿して同じプロセスをくり返し、かくして論文が出版されるには長い時間がかかります。査読は英語で「peer review(同僚による評価)」と呼ばれるように、研究者同士がボランティアで行うものです。論文は研究者コミュニティーの中でお互いが研究と評価を担うことで出版される仕組みになっていて、私たちバードリサーチのスタッフも日常的にいろんな学術誌の査読を引き受けています。話がそれますが、ときおり捏造論文が見つかってニュースを賑わすことがあります。査読は手法、分析、推論の欠陥をチェックするためのもので、研究資料を精査してデータの捏造までチェックするわけではありません。ですから、論文が出版されたあとで同じ現象が再現されないことなどでデータの捏造が露見する場合があるのです。
さて、このような従来の論文出版のプロセスを変えるような動きが起こりました。それが、受理されていない論文を公開する「プレプリントサーバー(pre-print server)」という仕組みで、初のプレプリントサーバーであるarXivは1991年に立ち上げられました。受理前の論文には間違えや不完全な部分があるのですが、それでもプレプリントサーバーで公開することには次のような利点があります。
1.誰が第一発見者か明確になる
論文の査読に時間がかかっているあいだに、別の研究者の同じ発見が出版されてしまうと、第一発見者の名誉を譲ることになります。物理学や生命化学では同じ現象を競って研究しているため、誰が先に見つけたかは重大な関心事です。論文が出版された順ではなく、プレプリントサーバーに登録した順に発見者が決まるのであれば公平でしょう。実験論文の場合は同じ方法で現象が再現できるか追試が行われるので、査読に代わって追試結果が信憑性を担保するという考えもあります。なお学術誌は未発表の発見を記した論文しか受け付けませんが、プレプリントサーバーに登録された論文は発表済とは見なさないという考え方が一般的です(学術誌の対応状況はWikipediaにまとめられています)。
2.出版される前に自分の研究を知ってもらえる
これまで研究者は自分の業績として受理された論文しか示せませんでした。特に就職先や助成金を求めている若い研究者にとっては、業績リストに載せられる論文が少ないことが悩みの種になります。プレプリント論文は学術誌に認められた業績ではありませんが、研究者の業績を調べる必要があるときにプレプリント論文をダウンロードして読んでもらえれば、その人の業績評価に役立ちます。論文の草稿を見せても同じではないかと思うかもしれませんが、上記の第一発見者をめぐる競争を考えると、プレプリントサーバー以外で研究を公表することは真似をされる危険をはらみます。
3.研究結果がすぐ利用できる
ITのように技術が日進月歩で進む分野では、時間がかかって論文が出たときには時代遅れということが起こりかねません。新しい知見をどんどん発表して自由に利用できるようにすることで急速に技術が進歩するため、研究者の気風としてもプレプリントサーバーで論文を公開することが好まれるようです。
現象の追試が容易ではなく、技術が日進月歩するほどでもない鳥類学や生態学ではプレプリントサーバーの利用は進んでいませんでしたが、科学界の変化の波は、ついに私にも及んできました。私が共著者になったツバメの論文がプレプリントサーバーで公開されたのです。
生物系プレプリントサーバー BioRxiv
この論文が登録されたBioRxivは、生物分野のプレプリントサーバーです。自分のアカウントを作成して論文のPDFファイルをアップロードすると、基本的なチェックが行われ(あまりに非科学的な内容だと却下されるようです)、48時間以内に公開されます。BioRxivのFAQページから要点を書き出しておきます。
- BioRxivへの登録と閲覧は無料です。
- 英語で書かれた論文しか登録できません。
- 学術誌へ投稿する前、または投稿と並行して論文を掲載できます。すでに受理された論文は掲載できません。
- 著作権はコンテンツの再利用をしやすいクリエイティブ・コモンズ・ライセンスのうち、引用元を表示することを義務づけるCC-BYを使用します。(この記事のように翻訳や図の引用を行うことや、全文を転載することも認められます)
- 論文のリバイズ版を掲載することはできますが、登録した論文を削除することはできません。
- 論文が学術誌に受理されれば、BioRxivのドキュメントに当該論文誌のへのリンクが表示されます(タイトルが変わっていると自動リンクされないので、自分で申告が必要です)。
- 論文には自由にコメントを付けられます。
- 論文にはdoiが付与されて引用できるようになります。
渡りの分かれ目がツバメの種分化を起こしている
それでは、BioRxivに登録された論文の概要を紹介しましょう。下記のリンクで全文をお読みいただけますので、ご意見があれば私までお知らせいただければ幸いです。
チベット高原を迂回して渡る位置に存在する、鳥類の交雑個体が分布する領域にある、種の交雑阻害境界線には渡りの分かれ目が存在している。
Elizabeth S.C. Scordato, Chris C.R. Smith, Georgy A. Semenov, Yu Liu, Matthew R. Wilkins, Wei Liang, Alexander Rubtsov, Gomboobaatar Sundev, Kazuo Koyama, Sheela P. Turbek, Michael B. Wunder, Craig A. Stricker, Rebecca J. Safran. 2019. Migratory divides coincide with species barriers across replicated avian hybrid zones above the Tibetan Plateau. bioRxiv: 698597. https://doi.org/10.1101/698597

写真1. 2014年に東京でツバメの捕獲をしました。右からElizabethさん、Chrisさん、神山。
この研究はElizabeth Scordatoさんたちが、ロシア西部からユーラシア大陸を横断して日本に至るまでの各地で集めたツバメのサンプルを分析したものです。私は2014年の春に日本を訪れた彼女と一緒に、東京と埼玉でツバメの捕獲を行いました(写真1)。
さて、この論文は生物の種分化が起きる要因についてのある仮説を検証したものです。生物の新種は、はじめに種から亜種が分化し、さらに亜種同士が完全に交配できないくらい違ってしまうことで生まれると考えられています。この種分化の要因のひとつとして「Migratory Devide」仮説が提唱されています。Migratory Devideとは越冬地が異なる渡り鳥が繁殖する場所の境界のことですが、定まった和訳がないので、この記事では「渡りの分かれ目」と呼ぶことにします。渡りの分かれ目では、異なる越冬地からやってくる個体が繁殖地に着くタイミングが違うので交配しにくいことや、交配した場合でも渡りの行動が遺伝的に決まっていると、生まれた子孫が正しい時期や、正しい方角へ渡れないことで生存率が低くなるために種分化が促進されるのではないか、と仮説では考えられています。

写真2. ツバメ3亜種は喉から腹の色に違いがある。左からH.r.rustica(イラン, 撮影 渡辺仁)、H.r. tytleri(ロシア・モンゴル国境, 撮影 照井滋晴、H.r. gutturalis(日本, 撮影 宮本桂)。
「渡りの分かれ目」は尾羽の炭素成分で分かる
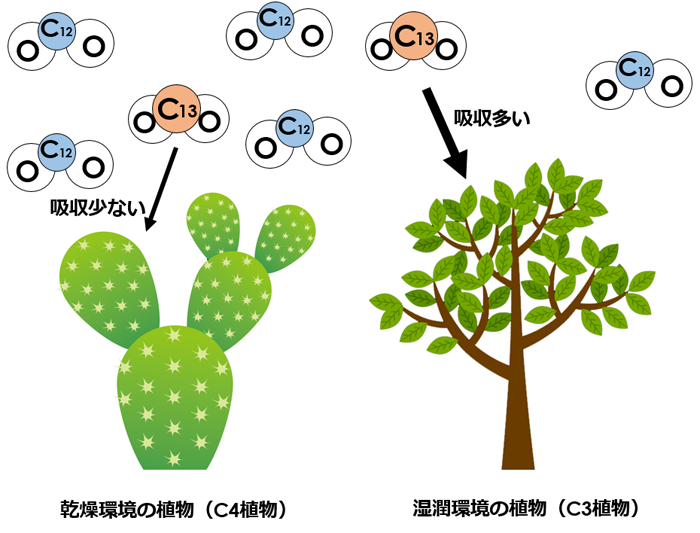
図2. 炭素同位体C12とC13からできている二酸化炭素が植物に吸収されるイメージ図。自然界の炭素の大部分はC12で、少量のC13が存在する。それぞれの炭素を含む二酸化炭素は質量が異なり、植物の光合成の仕組みの違いによって吸収される炭素同位体の比率に違いが生じる。
ツバメはヨーロッパから日本にかけて、Hirundo rustica rustica、H.r. tytleri、H.r. gutturalisの3亜種が分布しています(写真2)。学名のままでは分かりにくいので、便宜的に「ヨーロッパ亜種」「ロシア亜種」「アジア亜種」と呼ぶことにしましょう。まず渡りの分かれ目の存在を調べるために、ツバメの尾羽に含まれる炭素のうち同位体C13の割合を分析しました。炭素には質量が異なる同位体が存在し、相対的に重いC13をどのくらい吸収するかは植物によって異なります。そのため、食物連鎖の上位にいる動物の体を作る炭素の同位体比は、その地域の植物相に応じた特徴的な値を示します(図2)。この事実を使えば、ツバメの渡りを追跡しなくても大まかな越冬地を知ることができます。ツバメの尾羽は越冬地で生え替わるので、尾羽のタンパク質に含まれる炭素は越冬地の同位体割合を反映しているのです。尾羽の同位体分析の結果、東に分布するロシア亜種とアジア亜種ではC13の割合がほぼ同じで、東南アジアに特徴的な割合になっていました。一方、西に分布するヨーロッパ亜種のC13割合は、アフリカの乾燥地帯に特徴的な値であることがわかりました。つまり、図3の東経90~100度付近にあるヨーロッパ亜種-ロシア亜種の境界、そしてヨーロッパ亜種-アジア亜種の境界には渡りの分かれ目が存在しているというわけです。
DNA分析で亜種の交雑が起きている地理範囲が分かる
次に、捕獲したツバメのDNAから3亜種の遺伝子が地理的にどう広がっているかを分析しました。図3の地図では色が濃いほど亜種本来の遺伝子に近く、色が薄くなるにつれて他亜種と交雑している状態を示しています。3つの亜種は明確に別れているのではなく、ある亜種から別の亜種へ遷移する遺伝子のグラデーションが存在しているのです。
地図上の破線ABCはヨーロッパ亜種とロシア・アジアの2亜種の境界線を横切るツバメ捕獲ルート、点線DEFはロシア亜種とアジア種の境界線を横切るツバメ捕獲ルートです。両ルートに沿って遺伝子、尾羽のC13値、腹側の色が連続的に変化する様子を示したグラフを地図の左右に配置してあります。左側のヨーロッパ亜種からロシア・アジア亜種へ移り変わる3カ所のルートに沿った変化グラフでは、遺伝子の変化の中心と腹側の色の変化の中心は、尾羽の炭素同位体値の変化の中心、すなわち渡りの分かれ目と近い位置で起きています。ところが右側の3カ所のグラフでは、炭素同位体値の変化がほとんどない、つまり渡りの分かれ目がない状態で、遺伝子や腹の色の変化には左のグラフほど明確な変位点がありません。これらのことから、渡りの分かれ目がある場所では亜種の交雑が少なくなっていることが分かりました。
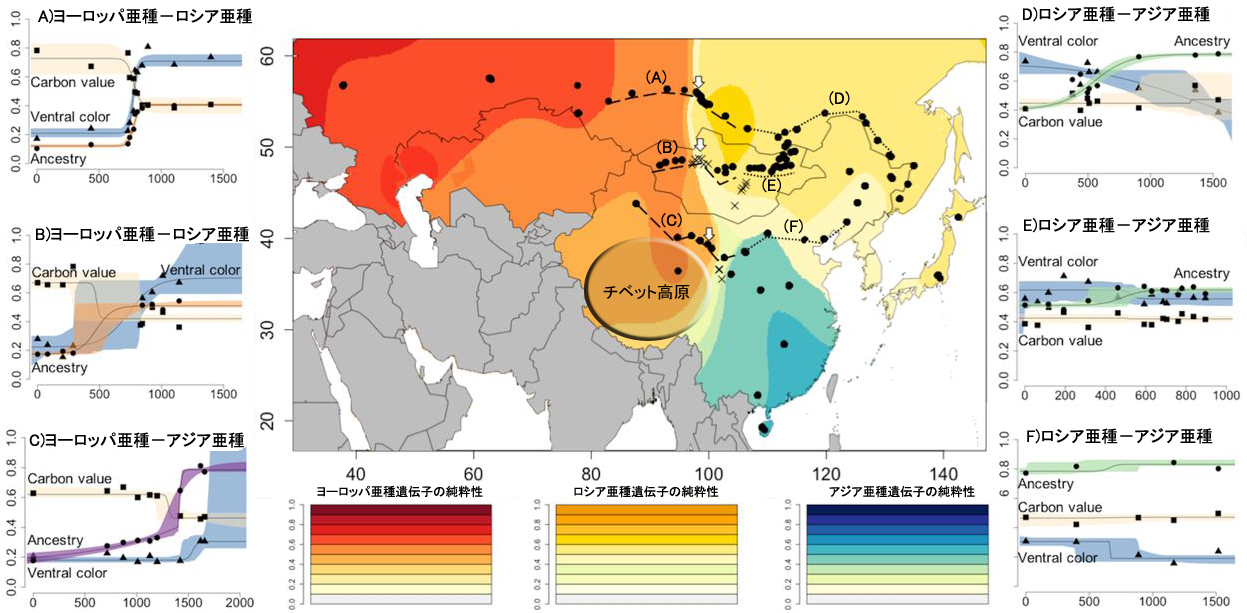
図3. ツバメ3亜種遺伝子の地理的分布(中央)、及び遺伝子・腹の色・越冬地を表す炭素同位体値の変移グラフ(左右)。地図の色が濃いほど亜種本来の遺伝子が純粋。グラフは横軸が捕獲ルートの西端からの距離、縦軸が0-1間で標準化した要素の値。各要素はCarbon value:炭素同位体値、Ventral color:腹の色、Ancestry:遺伝子の純粋性。論文の図にルートを示すアルファベットやチベット高原の位置を追記して作成した。
そのほかにも、この論文では、渡りの分かれ目が存在する亜種境界では、他方の亜種境界に比べて、遺伝的に近い雌雄でつがいになっていることなどが示されています。アジアからロシアにかけて繁殖する野鳥にはチベット高原を避けて東西方向に渡る種が少なくないので、このツバメの調査で分かった渡りの分かれ目と種分化の関係は、他の種にも適用できると考えられます。
日本の「アカハラツバメ」の正体
ところで話が論文の内容から離れますが、図3では日本で繁殖しているツバメはロシア亜種(tytleri)とアジア亜種(gutturalis)の中間的な個体であることが見て取れます。ロシア亜種のもっとも純粋な形質を持つツバメはモンゴル中央からロシアにかけての範囲に生息しており、この地域のツバメは日本鳥類目録第7版では亜種H.r. saturataとされています。この和名がアカハラツバメなのですが、この論文が準拠している最新のツバメの亜種分類では、ロシアからアジアにかけて生息するツバメは地理的分布による遺伝子の差に明確な境界がないため、アカハラツバメはロシア亜種の範疇に位置づけられています。日本で繁殖しているツバメには下腹部が赤い個体がときどき見られますし、越冬季には腹の赤い個体が多いとも言われています。これらはアカハラツバメが日本までやってきているのではないかとも考えられていたのですが、この論文のDNA解析によれば、腹の赤いロシア亜種と白いアジア亜種の雑種ということになります。腹の赤さの程度がさまざまなのは、雑種ツバメに遺伝子の多様性があるからなのでしょう。
2020年1月7日追記
この論文がEcology Lettersに掲載されました。https://doi.org/10.1111/ele.13420
