2017年度の調査研究支援プロジェクトでご支援いただいた澤田明と申します。その節はご支援いただきありがとうございました。改めて皆様にお礼申し上げます。調査研究プラン「南大東島に暮らす亜種ダイトウコノハズクの将来設計 産卵開始を急ぐことは有利な戦略か」で実施した研究を発展させてまとめた論文が、Population Ecology誌に掲載されましたので、その内容を紹介いたします。
私たちは南大東島に生息する亜種ダイトウコノハズクの長年にわたる調査データをもとに、統計的手法を用いて生存率や性比、産仔数、真の個体数など個体群動態に関する各種のパラメータの推定を行いました。その結果、2012年以降個体数が減少傾向にあること、そこにメスの生存率の低下が影響していることがわかりました。
紹介する論文
Akira Sawada, Tetsuya Iwasaki, Chitose Inoue, Kana Nakaoka, Takumi Nakanishi, Junpei Sawada, Narumi Aso, Syuya Nagai, Haruka Ono, Masaoki Takagi. 2021. Missing piece of top predator‐based conservation: Demographic analysis of an owl population on a remote subtropical island. Population Ecology.
https://doi.org/10.1002/1438-390X.12080
ダイトウコノハズク

図1 亜種ダイトウコノハズク。この個体は左足に黄色い足環がつけられていることで個体識別ができ、同一の巣を少なくとも6年使っていることがわかっています。
はじめに、研究材料となったリュウキュウコノハズクの亜種ダイトウコノハズクについて紹介します。亜種ダイトウコノハズクは沖縄県南大東島の固有亜種で、絶滅危惧Ⅱ類に指定されています。私の所属する研究グループは亜種ダイトウコノハズクの保全と学術研究を目的に、2002年から現在に至るまでこの鳥の繁殖モニタリングと標識再捕獲調査を続けています。1個体1個体を色のついた足環で識別することで、各個体の生存や産仔数、移動の履歴、血縁個体の居場所など、詳細な情報を集めてきました(図1)。亜種ダイトウコノハズクの個体群サイズは2005年の調査で210つがいとの見積もりがあります(Takagi et al. 2007)。
私たちはこの個体数調査以降、本亜種の保護のために島内に多数の巣箱を設置してきました。しかし、その後これまでに公式な個体数変化の解析はなされてきませんでした。適切に亜種ダイトウコノハズクの保護をすすめる上で、個体群に関する最新情報の整理が望まれていました。学術的調査と希少種保護の両方のために、私はこれまでの長年にわたる調査データを総合的に活用して、亜種ダイトウコノハズクの個体群動態(生存率や性比、産仔数、個体数、移出入率、繁殖個体割合などがどのように変化するのか)を解析することにしました。
調査研究支援プロジェクトで支援をうけた研究:生存率への着目
個体群動態研究の足掛かりとして、はじめに個体数の変動を規定する要因の一つである「生存率」と繁殖タイミングの関係についての研究を行いました。この研究は2017年の調査研究支援プロジェクトで支援を受けました。亜種ダイトウコノハズクは他の島の亜種リュウキュウコノハズクよりも繁殖時期が早いことが知られています。本亜種は縄張り性で、面積約30km2 (だいたい縦6km×横5km)の南大東島に高密度に縄張りを構えて生息しています。こうした環境資源の限られた状況では、繁殖を行うための縄張りをいち早く獲得することが子孫を残すうえでは重要です。
そこで私は「亜種ダイトウコノハズクの繁殖時期が早いのは、この島では早く繁殖するものほど適応的であるから(子孫を残すうえで有利であるから)」という仮説を立て、その検証を行いました。具体的には、統合個体群モデルと呼ばれる統計手法で個体群動態を解析し、繁殖時期が早いものほど生存率が高い傾向にあることを見出いしました(支援先 調査研究プラン 成果報告(2017年度)p.19-27 参照)。亜種ダイトウコノハズクは長生きするほど多くの子を残すことができるので(Sawada et al. 2020)、繁殖時期が早く長生きできる個体ほど多くの子孫を残せると考えられます。すなわち、はじめの仮説を支持する結果が得られました。
今回発表した論文の研究:個体群動態全体への着目
次に私は生存率に着目した研究を発展させて、個体群動態全体に着目した研究を行いました。2017年の調査研究支援プロジェクトの研究では生存率を調べるために統合個体群モデルという手法を用いました。実はこの手法は、生存率だけでなく性比や産仔数、個体数など様々な個体群統計量を推定することができます。なぜなら統合個体群モデルはそのモデルの中に、個体群動態を規定する複数のプロセス(生存や産仔、性比、個体数変化など)に対応した複数のサブモデルを含んでいるからです(Plard et al. 2019)。このモデルを表現する数式の部分は解析者が自由に改変することができるので、工夫次第で個体群動態に関する様々な統計量(生存率や性比、産仔数、個体数のほかにも移出入率、繁殖個体割合など)を求められるようになります。
2017年の調査研究支援プロジェクトの研究は亜種ダイトウコノハズクの個体群動態研究の第一段階だったので、シンプルな統合個体群モデルを用いました。例えば、性比はオスの割合が50%である(オス対メスが1対1である)とか、メスはすべて繁殖を行う、などの仮定をおき単純な数式で個体数変化をモデル化しました。しかし、今回発表した論文の研究ではそれらの仮定の多くを取り払い、より現実的な個体群動態の過程を扱えるように改良しました(図2)。例えば、性比はオスの割合がR %であるとか、メスのQ %が繁殖を行うと考え、RもQも仮定するのではなく推定するようにしました。今回発表した研究ではこうして作ったモデルに、2012年から2018年の間のデータを当てはめることで、個体群統計量を推定しました。それでは結果を示していきます。

図2 この研究で用いた統合個体群モデルの概略。四角で囲んだものがデータを表し、丸で囲んだものが推定すべき未知のパラメータを表します。データをもとにパラメータを推定します。サブモデルAは生存の記録データを未知のパラメータである個体の発見率と生存率で説明するモデル。サブモデルBはヒナの性別データを性比で説明するモデル。サブモデルCは巣立ちヒナ数データを産仔数で説明するモデル。サブモデルDは個体数の計数値(実際に島内を歩いて発見できた個体数)を発見率、生存率、性比、産仔数、真の個体数、ヒナの個体数、各性のヒナの個体数で説明するモデル。
統合個体群モデルによる推定結果
結果1:オスがメスよりも若干多い
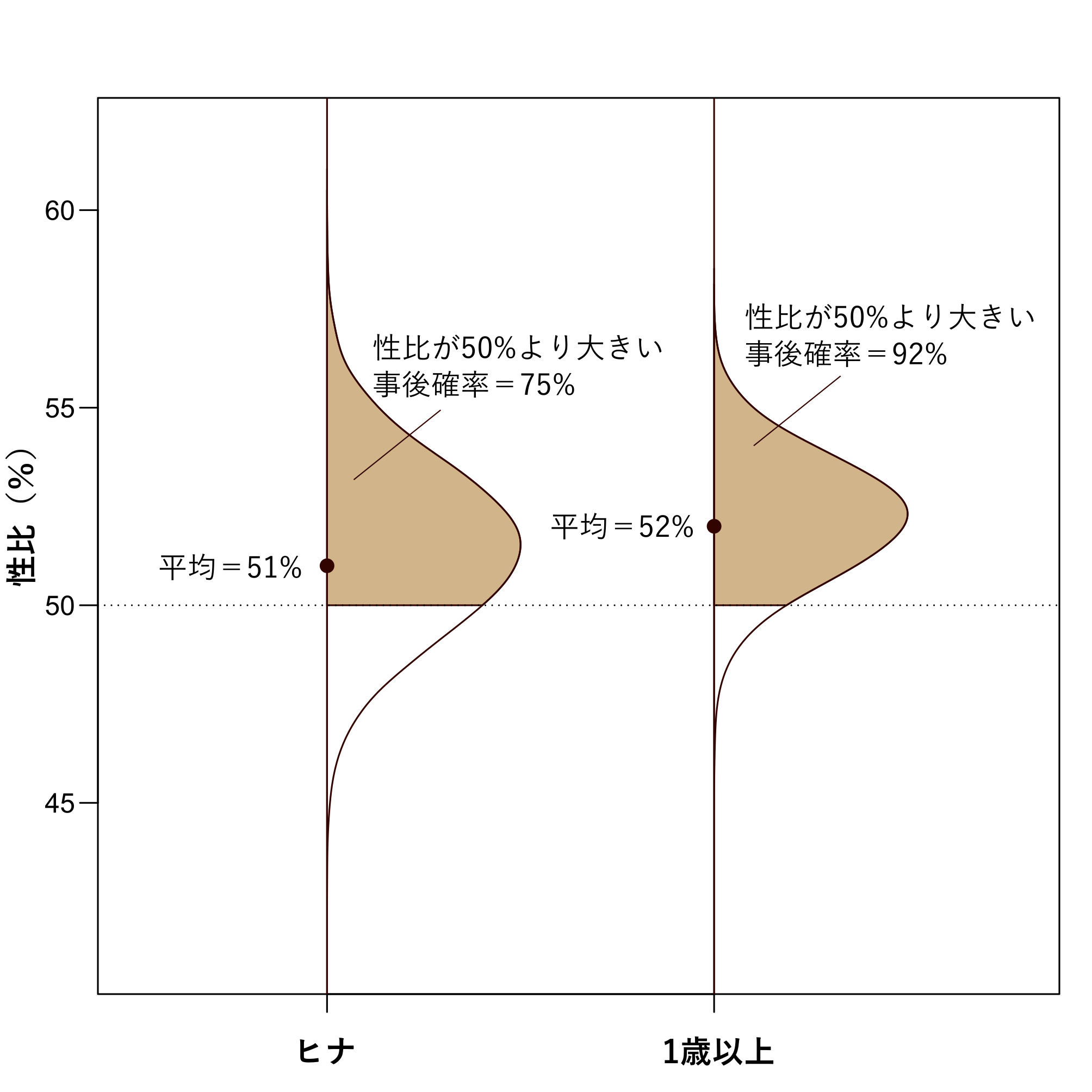
図3 ヒナと1歳以上の性比(オスの割合)の推定結果と性比の偏り。図中の横向きの山は性比がその値である確率の大小を表現している。例えば右の図からは、1歳以上の個体の性比は約52%である確率が高いと読み取ることができる。茶色の領域は性比が50%より大きい確率に対応する領域。
統合個体群モデルによる推定の結果、性比の推定値は、ヒナにおいてはオスの割合が51% (95%信用区間:47%-56%) で、1歳以上の個体についてはオスの割合が52% (95%信用区間:49%-55%)という結果になりました(図3)。ヒナから成鳥になるにかけてほんの数%ですがオスが多くなるという推定結果です。ここで、パラメータの95%信用区間とは、着目しているパラメータの値がその区間にある確率が95%であるという区間です。推定値の平均はわずかに50%からずれているだけですが、ヒナの性比がオスに偏る可能性は75%、1歳以上では92%と高い値になります。
結果2:成鳥の生存率は70%くらい
オス成鳥の年間生存率は74%、メス成鳥の年間生存率は73%と推定されました。有意な性差はみられないものの、メスの生存率はオスの生存率よりも1%小さいという結果になりました。
結果3:個体数はゆるやかな減少傾向
2012年から2018年にかけての個体数の推定値は、平均すると570.2個体でしたが、重要な結果として、個体数が減少傾向にあることがわかりました(図4)。個体群成長率の長期平均が0.98と推定されました。個体群成長率とは「(ある年の翌年の個体数)/(ある年の個体数)」で計算される値で1より小さい場合に個体数が減少していることを示します。
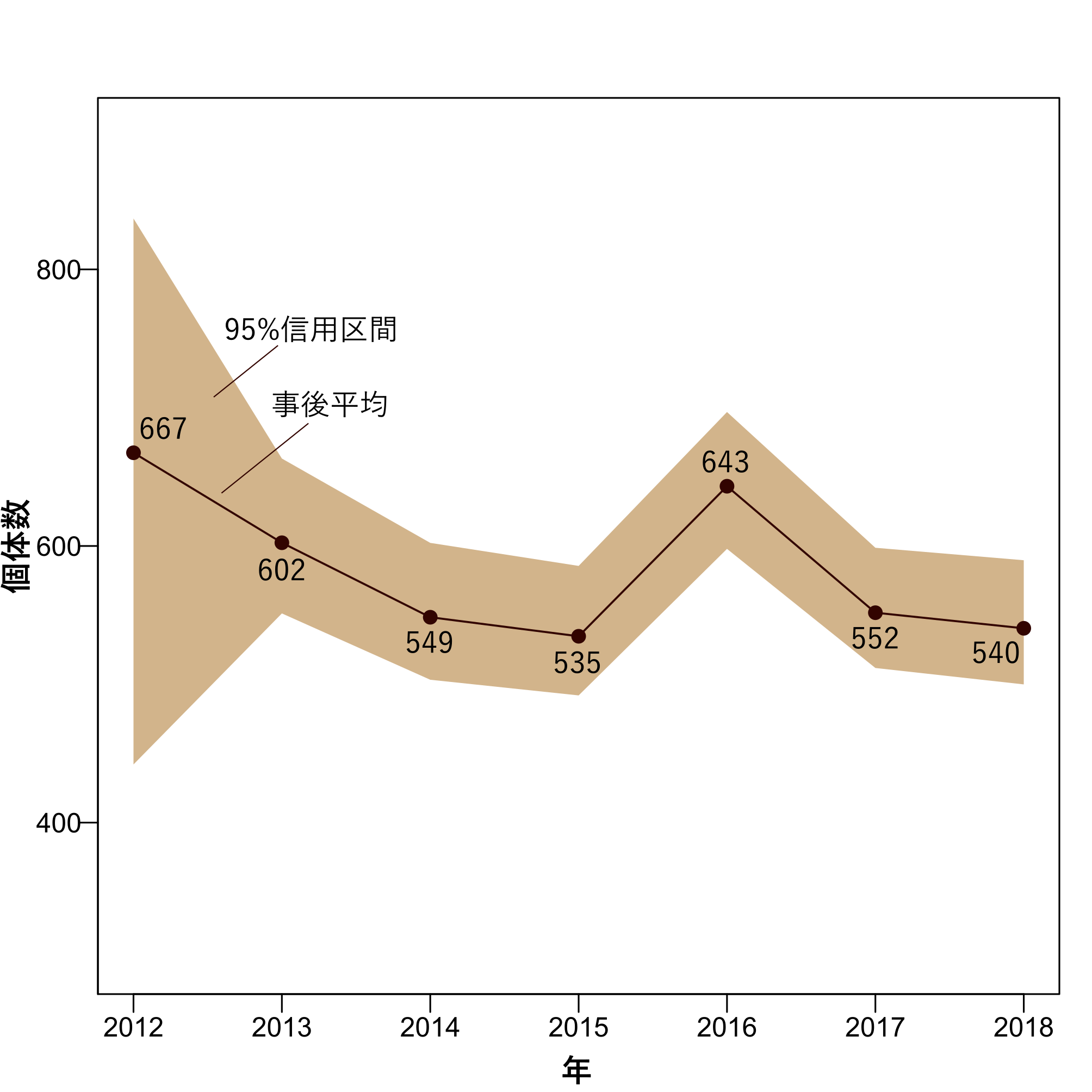
図4 個体数の推定値。実線と点が推定値の平均値を、茶色の領域は推定値の95%信用区間を示す。
結果4:メスの生存率の減少が個体数の減少をもたらす
今回の研究では統合個体群モデルの結果をさらにLife-stage simulation analysisという解析にかけました(Wisdom et al. 2000)。この解析をすることで個体群成長率が、他の個体群統計量(生存率や性比、出生率など)のどれに依存しているかを調べることができます。解析の結果、亜種ダイトウコノハズクの個体群成長率はメスの生存率ともっとも強い正の相関関係を示しました。したがって、メスの生存率の低下が亜種ダイトウコノハズクの個体数減少をもたらしている可能性が示唆されました。
ダイトウコノハズクの現状

図5 ネコによる捕食の事例。A、B:巣立ち目前の巣内ヒナが襲われた事例で、AとBは同一個体の写真。この個体は翼を奪われたのみで奇跡的に生き残って、現在も動物病院で保護さている。C:巣立ち前のヒナ2羽がすべて捕食された事例で、写真は巣の下に残っていた捕食痕。D:巣立ち後のヒナが捕食された事例。
今回推定された個体数570個体という数値は、2005年の調査で推定された210つがい(すなわち420個体)よりも大きい値になっています。巣箱設置がこの個体数の増加に貢献した可能性もありますが、個体数推定の方法が異なるため数値の単純な比較はできません。
その一方で2012年以降に関していえば個体数がわずかな減少傾向にあることがわかり、さらにメスの生存率がその減少傾向に影響していることが示唆されました。繁殖中のメスが巣内で卵やヒナを抱いているときに、卵やヒナごとネコに捕食されるという事例がしばしば確認されています(図5)。ダイトウコノハズクはメスのみが抱卵・抱雛するので、メスのほうがオスよりも捕食に会うリスクが高いのです。人為的に島に持ち込まれて野生化したネコによる捕食と、その捕食が特にメスに対して頻繁に生じることがメスの生存率低下をもたらし、個体群成長率を低下させているという可能性が考えられます。ネコによる捕食被害を詳細に調査する必要があります。
世界のフクロウ保護に貢献
フクロウ類は世界中に分布しており、研究も多くなされています。しかし、研究対象となっているのは温帯や大陸に分布する種なので、それらの種で得られた知見は熱帯や島嶼の種には当てはまらない可能性があります。例えば、Newton et al. (2016)は猛禽類の生存率に関する総説の中で、フクロウ類について体重から生存率を予測する回帰直線を示しています。体重86gの亜種ダイトウコノハズクの生存率は、この回帰直線をもとにすれば約57%と推定されますが、今回の研究ではそれより大きい73%から74%であることが示されました。亜熱帯海洋島の南大東島の比較的安定した環境に留鳥として生息していることがこの生存率の高さの原因のひとつと考えられます。
フクロウ類の多くの種は熱帯や島嶼に生息しているにもかかわらずそれらの研究は不足しています。特に個体群動態解析のような詳細なデータを必要とする研究はほぼ行われていません。特定の島に固有の種も多く、人知れずに絶滅の危機に瀕している種も多いと考えられます。生息地の減少やペットにするための捕獲などが彼らを脅かしていることは疑いようがありません。個体数を把握して管理するという保全活動のもっとも基礎的な部分で、個体群動態解析の結果は科学的な根拠を与えます。この研究は非温帯、島嶼域のフクロウ類の研究では類をみない精度で個体群動態の情報をまとめました。今後世界のフクロウ類の保護にも大きく貢献していくことが期待されます。
参考文献
Newton, I., M. J. Mcgrady, and M. K. Oli. 2016. A review of survival estimates for raptors and owls. Ibis. 158:227–248.
Plard, F., R. Fay, M. Kéry, A. Cohas, and M. Schaub. 2019. Integrated population models: powerful methods to embed individual processes in population dynamics models. Ecology 100:e02715.
Sawada, A., H. Ando, and M. Takagi. 2020. Evaluating the existence and benefit of major histocompatibility complex-based mate choice in an isolated owl population. J. Evol. Biol. 33:762–772.
Takagi, M., K. Akatani, S. Matsui, and A. Saito. 2007. Status of the Daito Scops Owl on Minami-daito Island, Japan. J. Raptor Res. 41:52–56.
Wisdom, M. J., L. S. Mills, and D. F. Doak. 2000. Life stage simulation analysis: estimating vital‐rate effects on population growth for conservation. Ecology 81:628–641.
