このシリーズでは毎回、研究者自身に、発表した英語論文について解説してもらいます。
今回は青木大輔さんに解説してもらいます。青木さんは北海道大学大学院理学院自然史科学専攻の博士前期課程2年生で、高木昌興教授の研究室に所属しています。今回解説してもらう論文は、青木さんが学部生のころに行ったカケスの系統地理についての研究で、2018年にドイツ鳥学会が発行するJournal of Ornithologyに掲載されました。この論文では、狭い日本海を挟んだ日本と東アジアの大陸の間で鳥がどのように種分化するかに注目しています。
【加藤貴大 編】
紹介する論文: Aoki D, Kinoshita G, Kryukov AP, Nishiumi I, Lee SI, Suzuki H (2018). Quaternary-related genetic differentiation and parallel population dynamics of the Eurasian Jay (Garrulus glandarius) in the circum-Japan Sea region. Journal of Ornithology 159: 1087-1097.
日本の固有種と種分化
みなさんにとって日本の固有種、と聞いてまず思いつく鳥は何でしょうか?ヤマドリ、アオゲラ、ルリカケス,アカコッコ,ノジコ・・・数えてみると案外多いな、いやいや他の国に比べれば少ないよ、いろんな意見があると思います。現在日本の固有種は全部で11種、日本でのみ繁殖する種を含めると17種が知られています。しかし、近年の目まぐるしいDNA解析の進展とともに、日本の鳥類の“本当の姿”を探る研究が少しずつ進んできています。今まで「種」は、主に形態の違いから判断されてきましたが、近年は行動や生態、そしてDNA情報の違いが注目されています。先行研究では、DNA情報に基づくと、同種だと思われていた日本列島とユーラシア大陸の多くの鳥で遺伝情報が大きく違っていることも分かっています。これをきっかけに、「実は日本には固有種が多数存在するかもしれない」と考えられるようになりました。
祖先種から2種が生まれる「種分化」の過程の多くは、ある地理的障壁(川や海、山など)が集団を2つに分けること(①)から始まります。もし2つの集団間で個体の移動、すなわち遺伝子の交流が地理的障壁によって妨げられた状態で十分な時間が経過すれば(②)、2集団は独自に進化します。やがて2集団は別の種になる(種分化する)のです。固有種が多い土地、例えばハワイ諸島を思い浮かべてみましょう。このような大海の孤島には何らかの原因で大陸や大きな島から偶然やって来る以外、鳥が集団を作ることはありません(①)。その頻度は非常に低いため、一度集団を作れば元の大陸集団とは交流は断絶されます(②)。なるほど、大陸など鳥がたくさんいる土地から距離が離れれば種分化が起きやすく、固有種が多くなるのは当然

図1.北海道に生息する亜種ミヤマカケス.最終氷期後に大陸から北海道に渡って来た可能性が示唆された.
でしょう。しかし、日本は島国とは言え、日本海は狭く、九州は対馬海峡、北海道は宗谷海峡を挟んですぐ大陸です。なぜ狭い日本海を挟んで固有種(候補)がそれほどまで多いのでしょうか?この謎を解き明かすことは、日本の鳥類の進化がどのように起こったかを理解できるきっかけになるため非常に重要な課題です。また、大陸に近い島での鳥の進化には謎が多く、そのプロセスを垣間見られる可能性も秘めています。
私はDNAを使って種分化のための道である①と②を日本の固有亜種で調べることにしました。これに適していたのが、カケス(Garrulus glandarius)でした。先行研究から、本州から九州に分布する亜種カケスG. g. japonicusは、ユーラシア北東部に分布する亜種ミヤマカケスG. g. brandtii (図1) や西ユーラシアに分布するG. g. glandariusとは、種に相当するDNA情報に違いがあることが分かっていました。また、カケスは海上を飛ぶことを嫌い、渡り途中でも海に来ると引き返す習性が知られています。陸地を介した移動が過去で推測しやすく、時代と共にカケスがどこを移動して来たかが分かりやすいわけです。
DNAからカケスが(亜)種分化した時期を推定
DNA情報を使うことで、私たちは進化の歴史を知ることができます。まず①いつどのような地理的障壁がカケスを大陸と日本列島の集団に分けたのかを調べました。分子時計という手法を使うことで、集団が分かれた年代(分岐年代)を調べ、その時代に何が起こったのかを推察しました(ミトコンドリアDNAの一部1、700塩基対を用いた)。すると、種カケス全体の祖先から一番初めに枝分かれしたのが日本の亜種カケスであり、この時代が約200万年前であることが分かりました。その後大陸の亜種が随時枝分かれしていったことも分かりました(図2)。
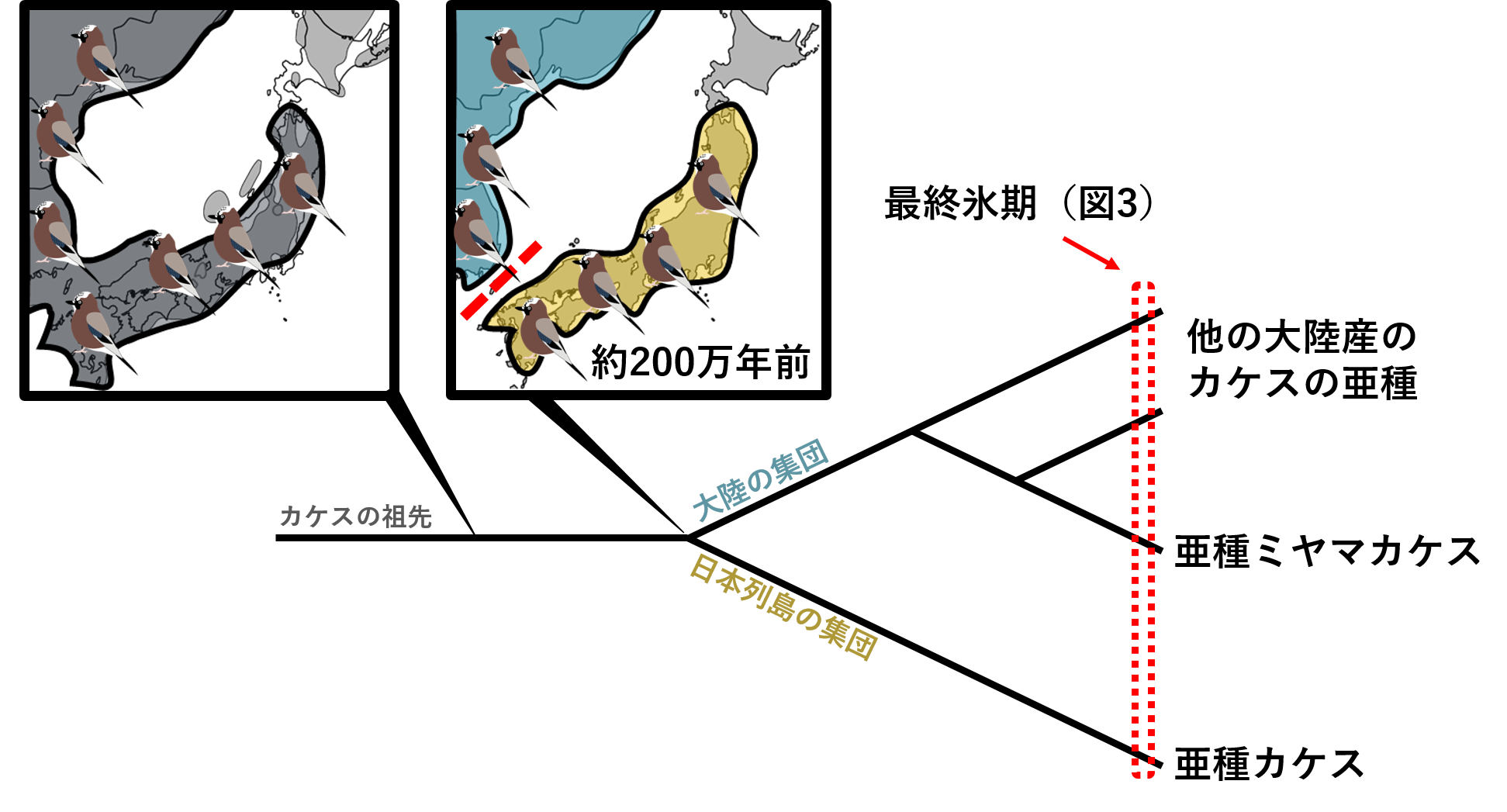
図2.日本列島と大陸のカケスの進化的関係(系統樹).2集団が分かれたのは丁度日本列島が列島になった時代と一致する.
約200万年前、時代は温暖な新第三紀から徐々に寒冷化し始め第四紀に突入した頃だといわれています。第四紀は氷河期と温暖期を激しく繰り返した時代です。水が氷として陸に存在する氷河期には海水面が低下し、温暖期には氷河は海へ流れ出るため、海水面が上昇していました。第三紀以前、日本列島は韓国・中国と陸続きの「半島」でした。しかし第四紀以降、海水面の上昇・低下により氷河期には半島に、温暖期には列島になったのです。カケスは陸地移動が主だと考えられますので第四紀が始まり、日本列島が半島から列島になった時、大陸から日本列島まで分布していたカケスの祖先は海で遮られ、日本と大陸で別の進化を歩み始めたと推測できます(図2)。実は200万年前には多くの鳥が日本列島と大陸で集団が分岐したと考えられています。例えば、メボソムシクイとコムシクイ、アオジとシベリアアオジ、クロジとシロハラホオジロなどです。これらの鳥はカケスと違いより長距離の渡りをする鳥です。つまり、カケスによって推測された進化のシナリオは、どうやら渡り鳥にも応用できそうです。
氷河期における大陸側と列島側の集団の分断
日本の亜種カケスは私が用いた亜種の中では一番古い時代から、独立に進化を歩み続けたわけですから、その間交流が遮断されている必要があります(②)。しかし、第四紀には何度も氷河期と温暖期を繰り返しており、列島と大陸が陸続きになる氷河期には再び集団間で交流が起こりそうなものです。なぜこのような交流は起こらなかった(起こりにくかった)のでしょうか?DNA情報からは、過去の集団の大きさや場所も推測できます。これによって、交流が遮断されていた理由が分かるかもしれません。
今回は最後の氷河期である最終氷期において、ユーラシア北東部(韓国―極東ロシア)の亜種ミヤマカケスと日本の亜種カケスを例に考えてみました。すると、寒く餌資源が少ない氷河期の間、大陸・列島では集団が小さくなり、比較的温暖な南方に避寒していたことが分かりました。大陸では極東ロシア南部の沿海州あたり、日本では西日本辺りがその避寒地(レフュージア)の候補地として推測されました(図3)。
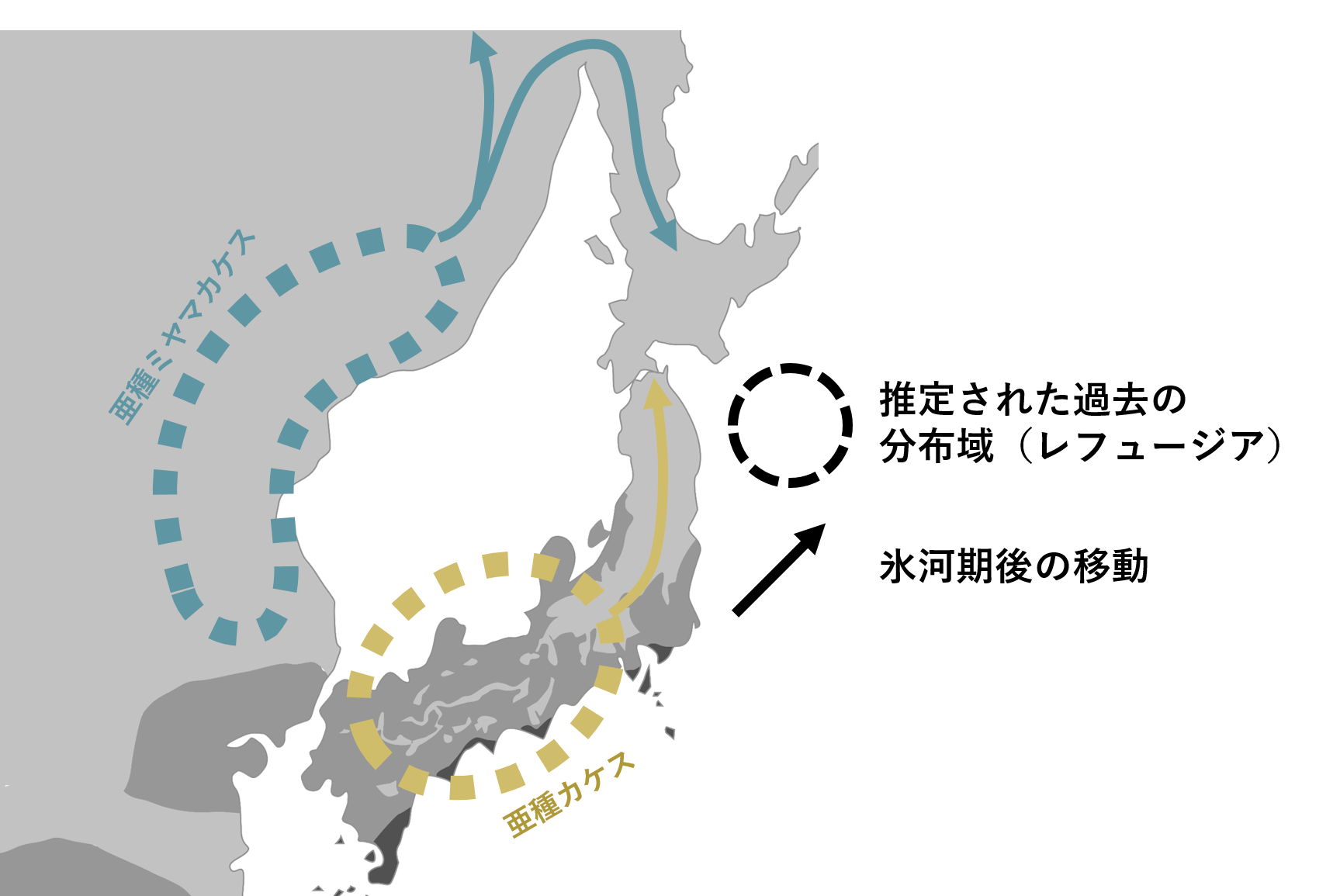
図3.最終氷期、列島と大陸は互いに繋がった(または非常に接近していた)が、それぞれの集団は互いに別の避寒地(レフュージア)に生息した.氷期後は分布を北に拡大したことでお互いが出会うことは無かったと考えられる.
その後徐々に暖かくなり現代のような温暖な気候になると、集団の大きさは大陸・列島でそれぞれ急激に大きくなり、分布を拡大し、現在の分布になったことが分かりました。つまり、氷河期の間陸続きだった列島と大陸ですが、集団が小さく互いに離れた地域に形成され、温暖期に集団は北へ分布を広げたことで、2集団間での交流が制限されていたのでしょう。 長距離を渡る鳥の一部でも列島と大陸の南方で集団が小さくなったと考えられています。渡り鳥でも同様な北上を氷期後経験したと考えると、渡り鳥でも列島―大陸間で種分化が引き起こされる理由を説明できるかもしれないと考えています。余談ですが、亜種ミヤマカケスは北海道やサハリンにも生息します(図1)。北海道やサハリンは氷河期に本州ではなく大陸と繋がっていました。先ほど説明した分布拡大の際にまだ大陸と繋がっていたサハリンと北海道では、カケスは北から移り住んだため、亜種カケスではなく亜種ミヤマカケスが住むことになったことがDNA解析から示されました。
東アジアとの国際的共同研究
DNAを詳しく解析することで、狭い日本海によって大陸から隔たれた日本列島で、固有種・亜種が生まれるメカニズムの一部が、私のカケスの研究から分かりました。従来、日本の鳥は、「大陸の鳥に上書きされたため歴史が浅い」と考えられてきましたが、日本は鳥類の進化に非常に興味深い土地だったのです。例え日本以外に分布を広げている種であっても、もしかしたら日本という土地がその鳥の進化に重要な機能を果たしたかもしれません。そのため、日本だけではなく、周辺諸国も併せて歴史をさかのぼる必要があります。今回の研究は私自ら海外に出向き、韓国・ロシアの研究者との共同研究を立ち上げました。歴史的混乱の多い東アジアですが、国の垣根を超えた研究が非常に重要になるのです。今回分かったのはカケスや東アジアの鳥の尤もらしい歴史のほんの一部でしょう。私は日本、韓国、中国、ロシアを含む「環日本海地域」で、カケスをはじめとした様々な鳥種の進化的背景を今後も探っていきたいと考えています。

著者紹介
青木大輔 学士(理学)
北海道大学大学院理学院 自然史科学専攻 修士2年
主にDNAを使った系統地理・集団遺伝学を用いて、東アジアにおける鳥類の種分化を研究している。現在はカケスの他、モズの生態進化も対象としている。
